
Lecture 16: Genetics and Evolution of Behavior
Contents
Lecture 16: Genetics and Evolution of Behavior¶
Genetics of Behavior¶
One general view in the study of the evolution of behavior is that behaviors can have a genetic basis. This is not to say that all behaviors are genetically based; indeed many behaviors are entirely culturally transmitted or learned and may have little to do with genetics (why are you attending college and not tending to the fields?). For genetically influenced behaviors we can treat them as we would treat any other genetically controlled trait of an organism: 1) if there are genetically based differences in a behavior, and 2) these differences affect fitness then, 3) behaviors can evolve by natural selection.
A few examples of behaviors with strong genetic components. Mating in Drosophila species is known to proceed according to stereotyped, set sequence of actions. Males court males in a very characteristic way which includes tapping, song singing, and tasting. Induced mutations in the fru (fruitless) locus break this system. Mutations in fru induce males to court others males in exactly as they would females. In fact this mutation is a sterile mutation- males will only court other males. Amazingly, because of sex-specific transcripts at fru, mutations which alter the female specific transcript also have strong effects on courtship. fru mutations have been recovered in which females court other females according to the very strict sequence that males normally use! This represents an incredible behavioral switch, which when rerouted will produce a complicated behavior under inappropriate circumstances (in some sense the behavior is “ectopic”). fru mutations are induced mutations though- what can we learn about natural variation from them? Probably not much.
Behavioral variation within populations¶
Natural genetic variation underlying behaviors does exist. Rover/sitter polymorphism is a great example. Among late instar larvae of Drosophila melanogaster two behavioral strategies are applied to foraging- there are rovers and there are sitters. Rovers are seen to forage more readily between patches of food and as well will move around within patches of food. Sitters, as their name implies, are most apt to sit in one place, are less likely to forage between patches of food, and forage shorter distances within patches. Both rovers and sitters are seen in natural populations at a frequency of about 70% rover and 30% sitter. \(F_2\) segregation of a cross between rovers and sitters suggests a single locus (for- foraging) with the \(for^R\) rover allele showing dominance to the sitter allele \(for^S\). Cloning of the \(for\) locus has shown it to encode a cGMP-dependent protein kinase (a PKG).
Even in humans we can find genetic variation underlying behavioral differences. Good example is novelty seeking behavior in people and its association to natural polymorphism in Dopamine D4 receptor (\(D4DR\)). Other examples include alcoholism, smoking, ear wiggling, handedness, tongue curling, etc.. Many people have claimed to find evidence for genetic differences in IQ. In particular during 1990s there was debate over a book by Herrnstein and Murray called the Bell Curve, which claimed such evidence. In particular they showed evidence that IQ was heritable and that there were differences in IQ among populations divided along ethnic lines (e.g. African americans, European americans)- huge flaw here, do you see it? Making claims about genetic variation in human behavior is a dangerous business because even if you do it correctly people don’t necessarily understand heritability and the environment.
Behavioral variation between populations¶
If additive variation underlying behavioral polymorphism exists within populations, we should expect that over time populations will diverge from one another over time due to the joint actions of selection, mutation, and drift. A good example might be to think about all of the behavioral adaptation that has accompanied bats’ transition to flight. Clearly just having wings isn’t enough- you need to be able to use them. A huge component of evolution along that lineage had to be behavioral. How good an example is human behavioral evolution since our common ancestor with chimpanzees? Behaviorally are we that different or is most of our behavior instead culturally (i.e. environmentally) determined?
Adaptive behavioral evolution undoubtedly occurs however. The leafy seadragon might be a good example here. Not only do you need to look like seaweed to be a good cryptic mimic, you also need to remain motionless. Another good example of an adaptive behavior that has been examined at the genetic level is Drosophila sechellia and its preference for its toxic host plant Morinda citrifolia. D. sechellia is a close relative of D. melanogaster that over time has evolved to be more or less dependent on a specific plant Morinda which is toxic to all of its close relatives. For example if you put D. melanogaster on the active component in Morinda, octonoic acid, it will die if under 15 minutes. D. sechellia does not die when place on octonoic acid. In fact D. sechellia actually prefers Morinda fruit as a place to mate and lay eggs. Both the resistance to octonoic acid and the preference for the fruit have been examined genetically, and strong QTL have been found using a hybrid breeding scheme back to D. simulans another close relative of D. sechellia and D. melanogaster.
There are also behavior constraints. Head scratching with the hind leg in amniotes (reptiles, birds, mammals; those w ith an amniotic sac). Most reach the hind leg over the fore limb to scratch the head; that birds and mammals do it suggests that this behavior may have a genetically programmed basis and has been inherited through much of higher vertebrate evolution.
Evolution of behavior¶
Behavior is usually dissected into two components for analysis: Proximate causes/questions in which one asks how the behavior is performed and ultimate causes/questions in which one asks why the behavior is performed. Tinbergen has identified four questions to pose when analyzing a behavior 1) what is the cause, 2) what is the development (ontogeny), 3) what is the current function 4) what is the phylogenetic history. A strict course on evolution focuses more on the latter two questions (recall adaptation/preadaptation/exaptation discussion and the identification of current utility vs. historical origin).
Herring gulls breed is large colonies on the ground and defend territories. Two separate calls used for 1) advertising nest site (“choking” call) and 2) as a territorial claim (the “oblique pose” and “long call”). The Kittiwake also breeds in colonies but nests on vertical cliffs and its nest pad is its territory and breeding site. In this species only one behavior serves both functions: “choking” behavior is both defensive and part of mate recognition/pair formation. This is seen as an adaptive behavioral shift with respect to the nest location (steep cliff).
There are many behaviors that at first appearance do not seem “adaptive”. Infanticide in lions was first viewed as “aberrant” behavior by abnormal individuals because it was not “good for the species” (male lions displace other males from groups of females and their offspring, and frequently kill the cubs). It is true that killing infants is not, in the short term, an effective means of increasing population numbers of a species. BUT, we now know (post W.D. Hamilton’s 1963, 1964 papers on inclusive fitness and kin selection and G. C. Williams book on Adaptation and Natural Selection) that the more appropriate way to address such problems is to think about them in the context of whether the behavior is good for the individual.
In analyzing infanticide from the perspective of gene thinking it is 1) not adaptive for a male lion to invest reproductive effort in an individual with whom he shares no genes and 2) once the infant is killed it is advantageous for the female to come into estrous and have more offspring with the new male (this will increase her reproductive output over leaving with the displaced male, and not benefiting from other advantages of group living: foraging, avoiding predation on young). Given the situation for both male and female, the observed behaviors make sense in terms of propagating ones genes.
The role of the gene (or genes!) as the unit that is relevant in the evolution play an important part in two influential books in the mid 1970s Sociobiology by E. O. Wilson, and The Selfish Gene by Richard Dawkins. To grossly oversimplify one of their main messages: “an organism is just DNAs way of making more DNA”
If we take the case of bird migration we want to know how the bird navigates to the breeding location (solar and magnetic cues during flight), how the bird knows when to begin migration (internal clocks and changes in day length [physiological changes]). There is usually a high cost associated with migration so we also want to know why birds do it since many die in the process (more time for feeding, more available food). Individuals that do migrate must leave more offspring than those that do not - again gene thinking helps account for why the behavior exists
Modeling Behavior¶
Population genetic approaches to the evolution of traits rarely tell us why a phenotype affects fitness in a particular way; the models usually look at whether fitness increases or not. The optimality approach to the analysis of behavior attempts to builds models where different behaviors are treated as the traits and asks which one of these behaviors might evolve. The approach generally ignores the mechanics of underlying genetic basis of the behavior (i.e., its mendelian and transmission genetics). Optimal models assume there is a genetic basis and treat each behavior as a haploid (asexual) trait that is inherited intact.
While Gould and Lewontin (and many others) have criticized optimal models, the builders of optimal model (e.g., John Maynard-Smith) argue that the models do not assume that the organisms are optimal (because there are constraints on evolution of traits), but by treating the problem as an optimality issue, it can tell you what kinds of behaviors might evolve.
Two general type of optimal models: frequency independent models are designed independent of what other strategies are doing, and seek to define the conditions which might influence behavior (recall the “optimal foraging” model we described in the adaptation lecture where a bird assess, quality, availability, distance to food items, etc.).
Evolutionary Stable Strategies¶
Frequency dependent models are ones where the strategy of one type depends on the strategies and frequencies of other types in the population. The general approach is to look for Evolutionary Stable Strategies (ESS): a strategy that, if adopted by all, cannot be “invaded” by a mutant strategy. Here a strategy = the behavior of an individual in a certain situation. These models come out of a field of math called Game Theory and were originally developed during the cold war as ways to analyze what to do in case of a nuclear attack- seriously. They’ve also been used in economics, political science, and psychology, but perhaps there best use is in understanding how behaviors evolve.
In the simplest game we imagine that there are two players, \(A\) and \(B\), and that there interactions with each other as opponents will lead to different payoffs. We can then simply describe our payoff matrix as is seen in Fig. 15. In that figure \(a\) is the outcome to player \(A\) in an interaction with player \(A\), \(b\) is the outcome to player \(A\) in an interaction with player \(B\) and so on. In this generic setting we consider an ESS to be any strategy that when common cannot be invaded by any rare strategy. That occurs when \(a > c\) or when \(a = c\) but \(b > d\). In words this is when a strategy pays better against itself than against any other strategy playing against it.
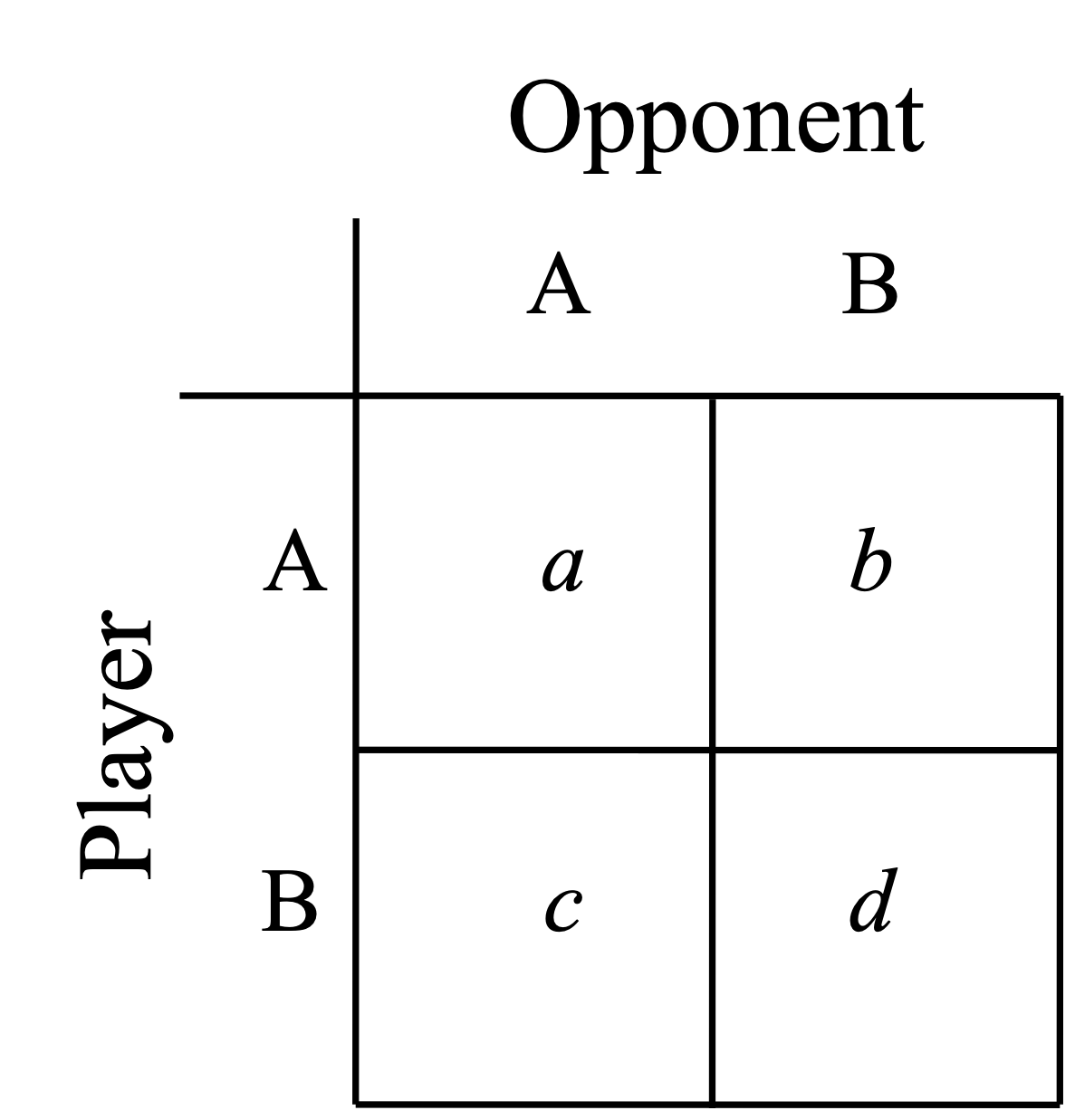
Fig. 15 Payoff Matrix from the simplest types of games¶
.
John Maynard-Smith was the first to apply game theory to evolution. He considered the case of ritualized aggression in species (like rams butting heads), and wondered how very few encounters led to death or serious injury. These types of model apply nicely to ritualized behaviors, distinct display behaviors which take the place of aggressive interactions. He considered a population composed of two strategies, hawks and doves. Hawks escalate fights and fight until they win or are defeated. Doves retreat from fights, but split a payoff if their opponents don’t fight. We now can fill in our payoff matrix and look for an ESS. The payoff matrix is shown below in Fig. 16.
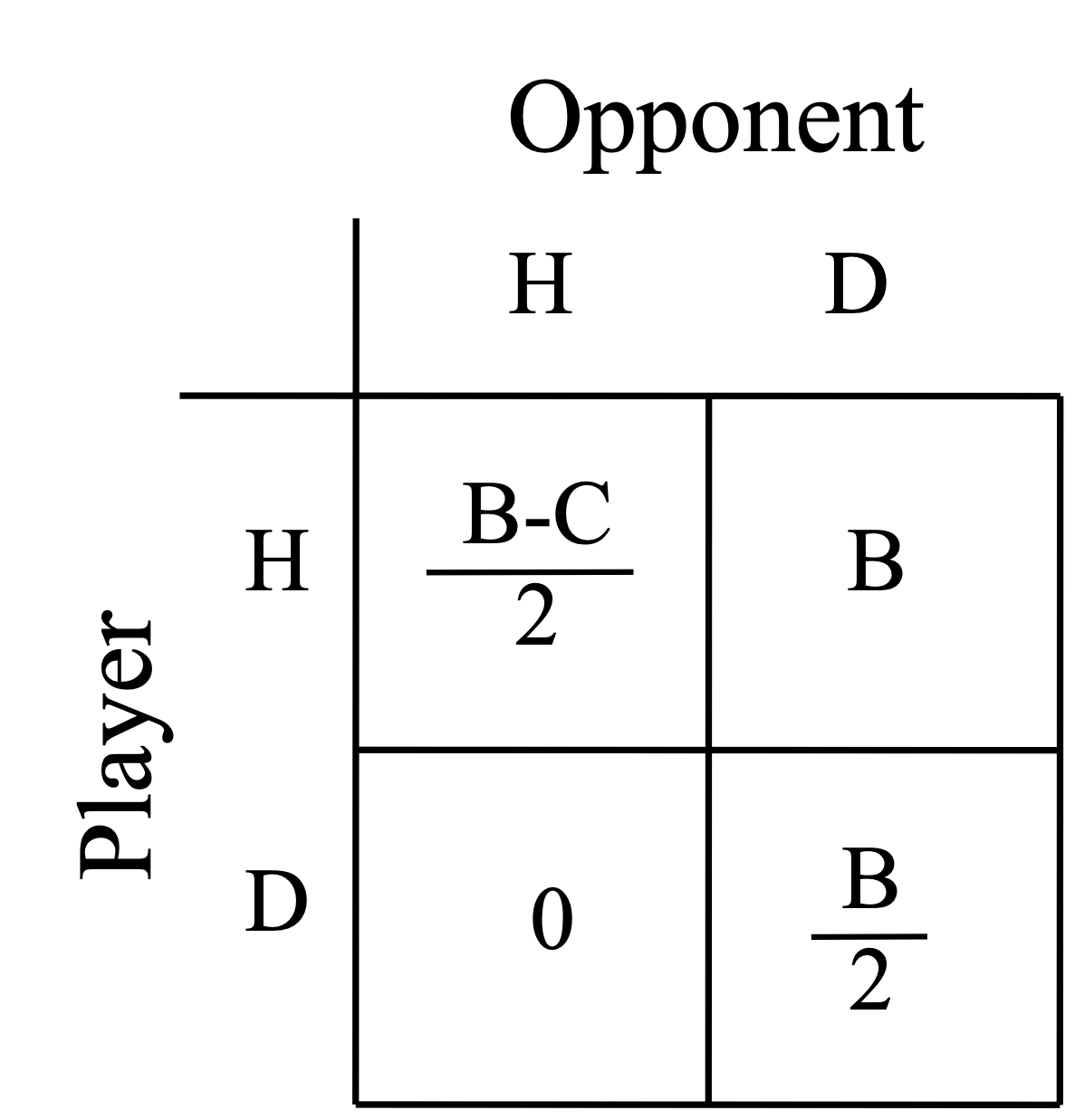
Fig. 16 Hawks and Doves Payoff Matrix B is the benefit of winning a contest. C is the cost of injury in a fight.¶
.
In a hawk:hawk interaction the payoff is \((B-C)/2\) because we assume that each hawk will win on average half the time. Dove:dove interaction yields \(B/2\) to player because payoff is split between the doves. Now lets look for ESS. If \(B>C\) ESS is being a hawk. What if \(B<C\)? No single ESS- population where hawk is common, dove strategy is better, in population where dove common, hawk is better. Population will evolve to have mixture of strategies where Freq(hawk) = B/C and Freq(dove) = 1 - B/C. So in essence the ESS is a mixed strategy where player should play two strategies with above probabilities.